

| 组别 | 一 | 二 | 三 | 四 |
| 温度/℃ | 27 | 28 | 29 | 30 |
| 光照后质量变化/mg | +4 | +5 | +6 | +6 |
| 暗处理后与光照前重量变化/mg | +3 | +3 | +3 | +1 |
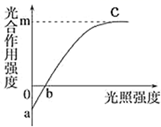
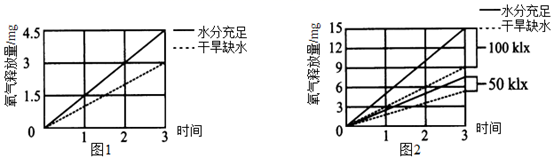
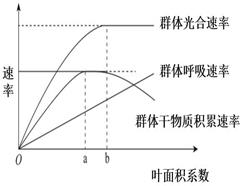




18∶00时叶肉细胞内产生ATP的细胞器有。
|
组别 |
处理[ABA浓度(μmol·L-1)] |
叶绿素含量(mg·g-1DW) |
净光合速率(μmolCO2·m-2·s-1) |
气孔导度(mmolH2O·m-2·s-1) |
胞间CO2浓度(μmol·m-2·s-1) |
|
T1 |
正常浇水+ABA(0) |
10 |
5 |
0.078 |
260 |
|
T2 |
正常浇水+ABA(200) |
6 |
5.8 |
0.051 |
200 |
|
T3 |
正常浇水+ABA(500) |
4 |
2.5 |
0.048 |
275 |
|
T4 |
中度干旱+ABA(0) |
12 |
1.2 |
0.031 |
300 |
|
T5 |
中度干旱+ABA(200) |
9 |
1.5 |
0.028 |
280 |
|
T6 |
中度干旱+ABA(500) |
7 |
1.3 |
0.039 |
320 |
|
T7 |
重度干旱+ABA(0) |
8 |
-1.6 |
0.041 |
450 |
|
T8 |
重度干旱+ABA(200) |
13 |
-1.3 |
0.043 |
425 |
|
T9 |
重度干旱+ABA(500) |
16 |
-0.8 |
0.068 |
405 |
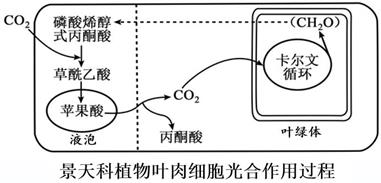
①植物形成糖类要经历卡尔文循环,卡尔文循环包括等过程。光反应为卡尔文循环提供。
②据图推测,图中细胞左侧的活动发生在(填白天或夜间),而右侧的活动发生在(填白天或夜间),作出这种推测的理由是。
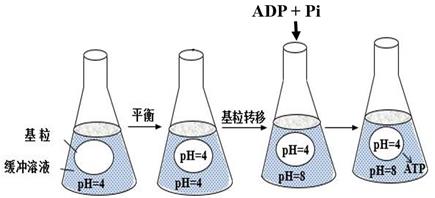
③为获取叶绿体,先要破碎叶肉细胞,再用法分离。
④上述实验结果表明,基粒类囊体合成ATP的原因可能是。据此推测,叶绿体在自然状态下产生ATP的过程中,光能的具体作用是。
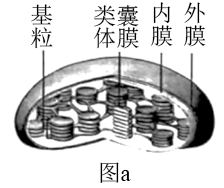

|
|
叶绿体A:双层膜结构完整 |
叶绿体B:双层膜局部受损,类囊体略有损伤 |
叶绿体C:双层膜瓦解,类囊体松散但未断裂 |
叶绿体D:所有膜结构解体破裂成颗粒或片段 |
|
实验一:以Fecy为电子受体时的放氧量 |
100 |
167.0 |
425.1 |
281.3 |
|
实验二:以DCIP为电子受体时的放氧量 |
100 |
106.7 |
471.1 |
109.6 |
注:Fecy具有亲水性,DCIP具有亲脂性。
据此分析:
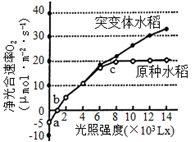
①叶绿体A和叶绿体B的实验结果表明,叶绿体双层膜对以(填“Fecy”或“DCIP”)为电子受体的光反应有明显阻碍作用,得出该结论的推理过程是。
②该实验中,光反应速率最高的是叶绿体C,表明在无双层膜阻碍、类囊体又松散的条件下,更有利于,从而提高光反应速率。
③以DCIP为电子受体进行实验,发现叶绿体A、B、C和D的ATP产生效率的相对值分别为1、0.66、0.58和0.41。结合图b对实验结果进行解释。


①图中,装置模拟的是发生在叶绿体基质中的反应;装置乙中的(填I ”或“ II ”)可表示C3。若光照强度增大,太阳能电池提供的电能增多,则短时间内I 的含量将(填“增加”或“减少”)。
②与植物体相比,固定等量的CO2时,装置乙输出的糖类的量多于植物积累的糖类的量,原因是 。

