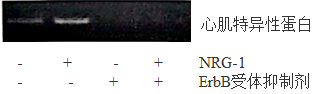
![]()


材料处理 | 实验结果 | |
| 叶表皮细胞状态 | 气孔开度 |
滴加蔗糖溶液 |
|
|
滴加清水 |
|
|

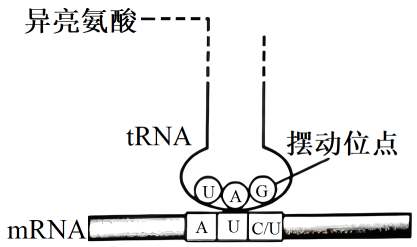
据图可知蓝光激活了H+—ATP酶,该酶发挥作用水解ATP,驱动H+以的方式运出保卫细胞,改变了细胞膜两侧电位,进而激活K+通道,K+内流导致细胞内渗透压,保卫细胞吸水,使气孔打开。
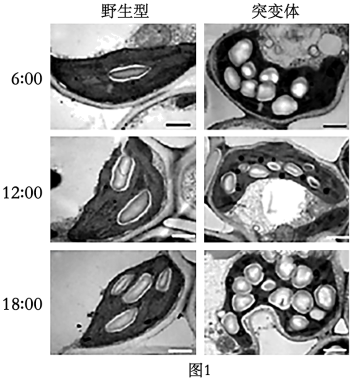
结果显示:突变体叶绿体中积累![]() 淀粉粒数目在三个时间点均野生型;随时间推移,野生型水稻的叶绿体中不断积累淀粉粒,原因是其叶绿体白天光合作用制造的有机物多于。
淀粉粒数目在三个时间点均野生型;随时间推移,野生型水稻的叶绿体中不断积累淀粉粒,原因是其叶绿体白天光合作用制造的有机物多于。
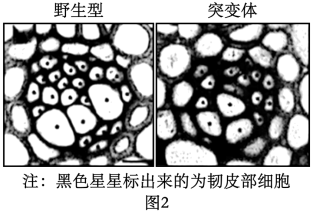

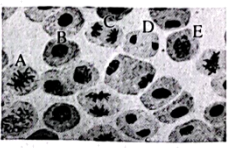
①图A中同源染色体两两配对的现象称为;配对的同源染色体的会发生染色体互换,导致同源染色体上的基因重组。
②图C中同源染色体会发生彼此分离,移向细胞两极的非同源染色体。
③图F中的细胞处于期;花粉母细胞经减数分裂最终形成的子细胞中染色体数目为体细胞的。若将四倍体三浅裂野牵牛的花粉进行离体培养,获得的植株称为。
核供体细胞 | 骨髓间充质干细胞 | 胎儿成纤维细胞 | |
供体细胞内RNA m6修饰水平 | 1.0 | 0.7 | |
重构胚的 发育效率 | 发育至卵裂期的重构胚占比 | 71.2% | 55.3% |
发育至囊胚期的重构胚占比 | 26.3% | 15.9% | |
结果表明核供体细胞的分化程度越高,。
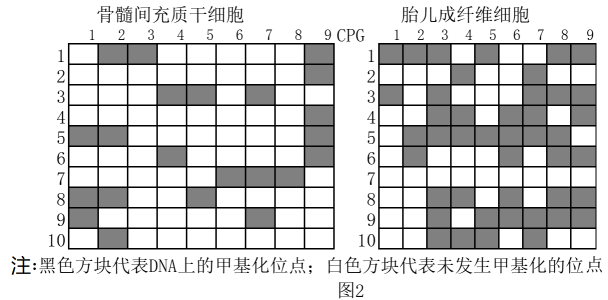
基因上游序列存在酶的结合位点,该酶可催化基因的转录。据图2可知成纤维细胞M基因上游序列的甲基化程度骨髓间充质干细胞。
欲设计实验验证M基因上游序列的甲基化程度是导致不同供体细胞RNA m6A修饰水平差异的原因。思路如下:
实验组用DNA甲基化抑制剂处理骨髓间充质干细胞,对照组不处理;检测两组的M基因上游序列甲基化水平。
请修正实验方案: 。

。



据图可知突变体eg的果实滞绿性状由基因控制,遗传遵循定律。对F1进行测交,检测后代果实色素含量,叶绿素含量显著高于野生型的后代占。叶绿素和番茄红素均不是蛋白质,推测上述基因是通过控制,控制色素的合成或分解,进而控制果实颜色。

据图可知,与野生型相比,突变体eg的SGR1基因碱基发生导致突变。根据突变位点前的测序结果可知,图中显示的DNA单链是SGR1基因转录的(填“模板链”或“非模板链”)。
真核生物基因初始转录产物(mRNA前体)在特定位点被识别后,部分序列被剪切掉,其余序列加工形成成熟mRNA,推测突变体egSGR1基因突变导致,最终引起编码产物肽链缩短。








