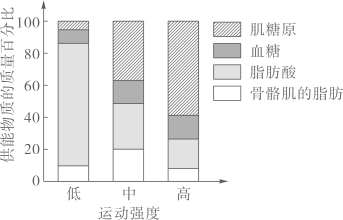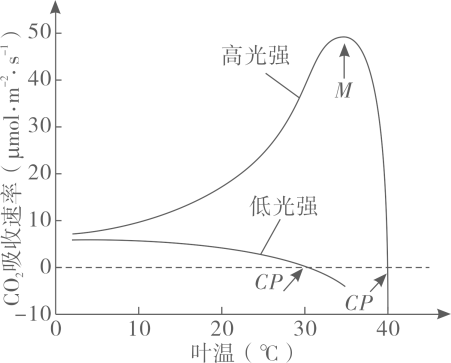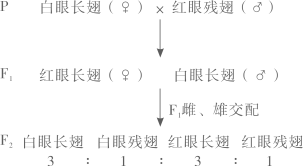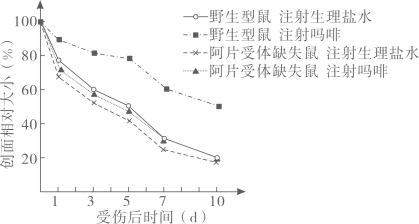
主要实验材料和设备:P菌、A菌、培养基、圆形滤纸小片、离心机和细菌培养箱。
①静息状态下,K+静电场强度为mV,与静息电位实测值接近,推测K+外流形成的静电场可能是构成静息电位的主要因素。
②为证明①中的推测,研究者梯度增加细胞外K+浓度并测量静息电位。如果所测静息电位的值,则可验证此假设。
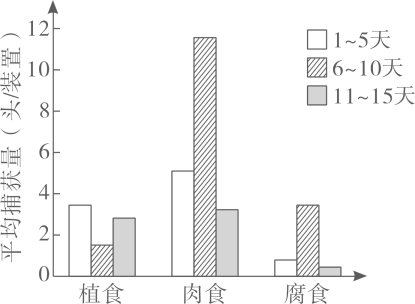
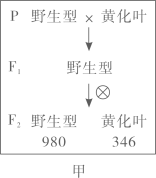
在大量繁殖A的过程中,会因其他品系花粉的污染而导致A不纯,进而影响种子S的纯度,导致油菜籽减产。油菜新生叶黄化表型易辨识,且对产量没有显著影响。科学家设想利用新生叶黄化性状来提高种子S的纯度。育种过程中首先通过一系列操作,获得了新生叶黄化的A1,利用黄化A1生产种子S的育种流程见图丙。

①图丙中,A植株的绿叶雄性不育子代与黄化A1杂交,筛选出的黄化A植株占子一代总数的比例约为。
②为减少因花粉污染导致的种子S纯度下降,简单易行的田间操作用。
调控植物细胞活性氧产生机制的新发现
能量代谢本质上是一系列氧化还原反应。在植物细胞中,线粒体和叶绿体是能量代谢的重要场所。叶绿体内氧化还原稳态的维持对叶绿体行使正常功能非常重要。在细胞的氧化还原反应过程中会有活性氧产生,活性氧可以调控细胞代谢,并与细胞凋亡有关。
我国科学家发现一个拟南芥突变体m(M基因突变为m基因),在受到长时间连续光照时,植株会出现因细胞凋亡而引起的叶片黄斑等表型。M基因编码叶绿体中催化脂肪酸合成的M酶。与野生型相比,突变体m中M酶活性下降,脂肪酸含量显著降低。
为探究M基因突变导致细胞凋亡的原因,研究人员以诱变剂处理突变体m,筛选不表现细胞凋亡,但仍保留m基因的突变株。通过对所获一系列突变体的详细解析,发现叶绿体中pMDH酶、线粒体中mMDH酶和线粒体内膜复合物I(催化有氧呼吸第三阶段的酶)等均参与细胞凋亡过程。由此揭示出一条活性氧产生的新途径(如图):A酸作为叶绿体中氧化还原平衡的调节物质,从叶绿体经细胞质基质进入到线粒体中,在mMDH酶的作用下产生NADH([H])和B酸,NADH被氧化会产生活性氧。活性氧超过一定水平后引发细胞凋亡。
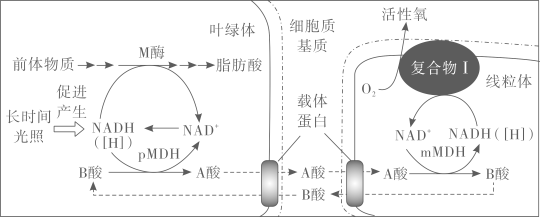
在上述研究中,科学家从拟南芥突变体m入手,揭示出在叶绿体和线粒体之间存在着一条A酸-B酸循环途径。对A酸-B酸循环的进一步研究,将为探索植物在不同环境胁迫下生长的调控机制提供新的思路。
①确定相应蛋白的细胞定位和功能
②用诱变剂处理突变体m
③鉴定相关基因
④筛选保留m基因但不表现凋亡的突变株
①纯合小鼠Lx:小鼠L基因两侧已插入特异DNA序列(x),但L的功能正常;
②Ce酶基因:源自噬菌体,其编码的酶进入细胞核后作用于x,导致两个x间的DNA片段丢失;
③Er基因:编码的Er蛋白位于细胞质,与Er蛋白相连的物质的定位由Er蛋白决定;
④口服药T:小分子化合物,可诱导Er蛋白进入细胞核。
请完善制备小鼠IK的技术路线:→连接到表达载体→转入小鼠Lx→筛选目标小鼠→→获得小鼠IK。