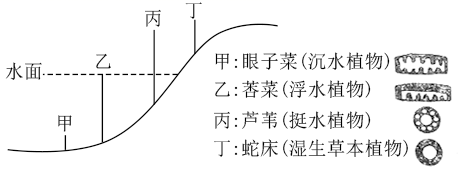
为提高转基因抗虫棉的抗虫持久性,可采取如下措施:
①基因策略:包括提高杀虫基因的表达量、向棉花中转入多种杀虫基因等。例如,早期种植的抗虫棉只转入了一种Bt毒蛋白基因,抗虫机制比较单一;现在经常将两种或两种以上Bt基同时转入棉花。
②田间策略:主要是为棉铃虫提供底护所。例如我国新疆棉区,在转基因棉田周围种植一定面积的非转基因棉花,为棉铃虫提供专门的庇护所:长江、黄河流域棉区多采用将转基因抗虫棉与高粱和玉米等其他棉铃虫寄主作物混作的方式,为棉铃虫提供天然的庇护所。
③国家宏观调控政策:如实施分区种植管理等。
|
个体 |
母亲 |
父亲 |
姐姐 |
患者 |
|
表现型 |
正常 |
正常 |
正常 |
患病 |
|
SC基因测序结果 |
[605G/A] |
[731A/G] |
[605G/G];[731A/A] |
? |
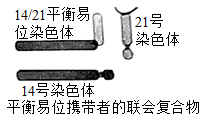
注:测序结果只给出基一条链(编码链)的碱基序列[605G/A]示两条同源染色体上SC基因编码链的第605位碱基分别为G和A,其他类似。
若患者的姐姐两条同源染色体上SC基因编码链的第605和731位碱基可表示为下图1,根据调查结果,推断该患者相应位点的碱基应为( )

 B .
B .  C .
C .  D .
D . 
为了研究疟原虫对青蒿素的抗药性机制,将一种青蒿素敏感(S型)的疟原虫品种分成两组:一组逐渐增加青蒿素的浓度,连续培养若干代,获得具有抗药性(R型)的甲群体,另一组为乙群体(对照组)。对甲和乙两群体进行基因组测序,发现在甲群体中发生的9个碱基突变在乙群体中均未发生,这些突变发生在9个基因的编码序列上,其中7个基因编码的氨基酸序列发生了改变。
为确定7个突变基因与青蒿素抗药性的关联性,现从不同病身上获取若干疟原虫样本,检测疟原虫对青蒿素的抗药性(与存活率正相关)并测序,以S型疟原虫为对照,与对照的基因序列相同的设为野生型“+”,不同的设为突变型“-”。部分样本的结果如表。
|
疟原虫 |
存活率(%) |
基因1 |
基因2 |
基因3 |
基因4 |
基因5 |
基因6 |
基因7 |
|
对照 |
0.04 |
+ |
+ |
+ |
+ |
+ |
+ |
+ |
|
1 |
0.2 |
+ |
+ |
+ |
+ |
+ |
+ |
- |
|
2 |
3.8 |
+ |
+ |
+ |
- |
+ |
+ |
- |
|
3 |
5.8 |
+ |
+ |
+ |
- |
- |
+ |
- |
|
4 |
23. 1 |
+ |
+ |
+ |
+ |
- |
- |
- |
|
5 |
27.2 |
+ |
+ |
+ |
+ |
- |
- |
- |
|
6 |
27.3 |
+ |
+ |
+ |
- |
+ |
- |
- |
|
7 |
28.9 |
+ |
+ |
+ |
- |
- |
- |
- |
|
8 |
31.3 |
+ |
+ |
+ |
+ |
- |
- |
- |
|
9 |
58.0 |
+ |
+ |
+ |
- |
+ |
- |
- |
回答下列问题:
|
实验方案 |
预期结果 |
|
I.转基因玉米×野生型玉米 II.转基因玉米×甲品系 III.转基因玉米自交 IV.野生型玉米×甲品系 |
①正常籽粒:干瘪籽粒≈1:1 ②正常籽粒:干瘪籽粒≈3:1 ③正常籽粒:干瘪籽粒≈7:1 ④正常籽粒:干瘪籽粒≈15:1 |
统计干瘪籽粒(F2)的数量,发现类型1最多、类型2较少、类型3极少。请解释类型3数量极少的原因。
|
密码子序号 |
1…4…19 20…540 |
密码子表(部分): |
|
正常核苷酸序列 |
AUG…AAC…ACU UUA…UAG |
AUG:甲硫氨酸,起始密码子 |
|
突变①↓ |
||
|
突变后核苷酸序列 |
AUG…AAC…ACC UUA…UAG |
AAC:天冬酰胺 |
|
正常核苷酸序列 |
AUG…AAC…ACU UUA…UAG |
ACU、ACC:苏氨酸 |
|
突变②↓ |
||
|
突变后核苷酸序列 |
AUG…AAA…ACU UUA…UAG |
UUA:亮氨酸 |
|
正常核苷酸序列 |
AUG…AAC…ACU UUA…UAG |
AAA:赖氨酸 |
|
突变③↓ |
UAG、UGA:终止密码子 |
|
|
突变后核苷酸序列 |
AUG…AAC…ACU UGA…UAG |
…表示省略的、没有变化的碱基 |

回答下列问题: